 ×
× 


以下文章来源于中国化妆品 ,作者卢云宇
编者荐语:
转载彗搏科技发表于《中国化妆品》杂志(China Cosmetics Review)(月刊)2021年3月刊的约稿文章~中国化妆品创刊于1992年,是中国商业联合会主管的国内外公开发行的国内唯一一本隶属一级单位直接管理的国家级权威中央媒体刊物~

精准护肤理念是以生物学靶点为依托,强调实验论证与人体功效之间的科学关联。线粒体,是真核细胞的生物能量学中心,是在能量生成和氧化应激中起首要作用的细胞器。线粒体不仅提供细胞90%以上的能量来源,而且深度调控能量代谢、ROS与自由基、氧化应激、炎症、伤口愈合、自噬、钙稳态、干细胞、组织重建、色素沉着、生物节律和毛发生长等功能。另外,线粒体被认为是细胞⁄机体衰老的主要原因,线粒体功能障碍导致皮肤老化与损伤,而靶向线粒体有助于恢复皮肤活力,精准解决深层修复抗衰难题。
线粒体于1857年被瑞士科学家Kolliker在肌肉细胞中发现,随后德国科学家Altmann将其命名为bioblast(原生粒)。1898年德国细胞学家Carl Benda依据其结构(希腊语Mitos线和Chondros颗粒)将其命名为mitochondrion(线粒体)。就学术研究领域而言,线粒体堪称诺贝尔奖专业户,自1931年至2019年,关于线粒体呼吸酶、三羧酸循环、呼吸链细胞色素氧化酶、三磷酸腺苷(ATP)合成、化学渗透理论、能量转换及ATP合成酶催化机理、细胞与线粒体自噬、细胞感知及缺氧诱导调控等线粒体相关研究频频获奖,且热度不减。
线粒体结构与功能
线粒体通常分布在能量需求旺盛的区域,是一种双膜结合体。学界已经证实,线粒体由好氧菌与原始真核细胞共生演化而来。这种进化史反映在线粒体DNA(mtDNA)的存在及其独特性上——一个具有母系遗传自主基因组的细胞器。mtDNA是双链超螺旋环状分子,是独立于核DNA外的唯一DNA,具有多拷贝性、有阈值效应、高突变性等特点,因此线粒体被称为半自主细胞器。mtDNA包含呼吸功能(复合体I、III、IV和复合体V)所必需的37个基因,其中包括编码13种氧化磷酸化(OXPHOS)蛋白、22种tRNA和2种rRNA。
线粒体与皮肤衰老
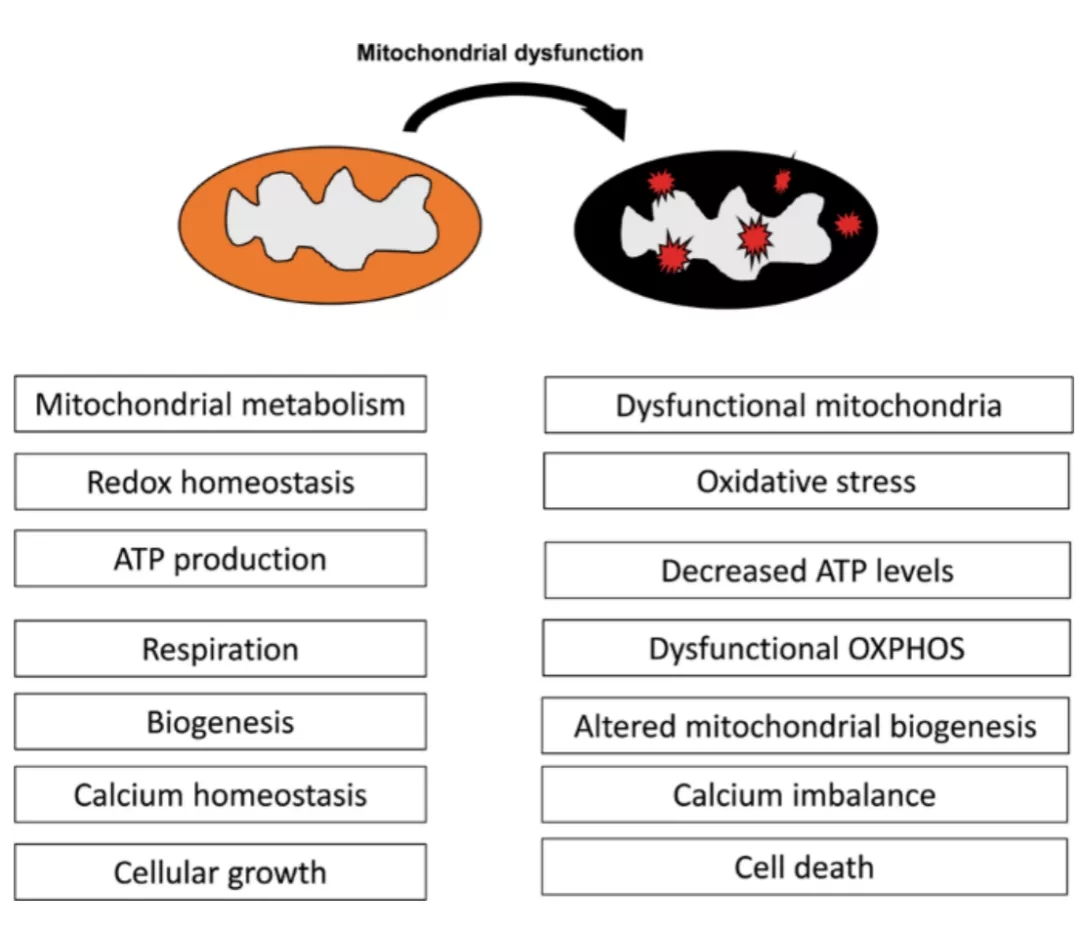
线粒体功能障碍与皮肤衰老
皮肤衰老分为年龄增长造成的的内源性衰老和日晒、环境污染、吸烟、生活方式等因素所致的外源性衰老,其关键特征是线粒体功能障碍的直接后果,这些衰老表型包括皱纹、头发变白和脱落、色素沉着不均以及伤口愈合缓慢等。在分子水平上,衰老皮肤的特征是真皮层和表皮层中同时存在的膜电位丧失、ROS水平升高、线粒体受损、导致核和mtDNA突变的DNA损伤、由于酶改变引起的呼吸链缺陷、细胞调节改变和疾病进展等。美国科学家哈曼(Harman)在20世纪50年代提出自由基损伤理论,认为线粒体损伤及功能障碍是衰老的始发环节,高水平ROS具有细胞毒性,作为能量代谢副产物影响线粒体功能,造成脂质、蛋白质、DNA损伤,而线粒体质量、活性改变及电子传递链复合体的生物合成直接影响细胞衰老。
线粒体呼吸、能量代谢
与电子传递链

线粒体结构
线粒体是真核生物的“能量工厂”,以及糖类、脂肪和氨基酸氧化释放能量的场所。有氧呼吸的核心环节——三羧酸循环在线粒体基质中完成,通过氧化磷酸化利用期间生成的还原型烟酰胺腺嘌呤二核苷酸(NADH)和还原型黄素腺嘌呤二核苷酸(FADH2)等高能分子转移电子与质子,积累电化学势差,还原氧气,制造三磷酸腺苷(ATP)。ATP是细胞的直接能量来源,含有高能磷酸键,极不稳定。线粒体能够检测并响应氧气水平,低氧被感应后可以引发一系列适应性反应,并激活低氧诱导因子(HIF)。如线粒体复合物III的泛醌循环可响应缺氧而产生活性氧(ROS),从而稳定HIF alpha蛋白。

线粒体与电子传递链
电子传递链(ETC)也被称为呼吸链,指线粒体在有氧呼吸过程中的电子转移反应,主要由位于线粒体内膜上的4种酶复合物I(NADH脱氢酶)、II(琥珀酸脱氢酶)、III(细胞色素c还原酶)、IV(细胞色素c氧化酶)和2种可移动电子载体CoQ(又称泛醌UQ)及细胞色素c构成。
线粒体与ROS
自由基及氧化应激
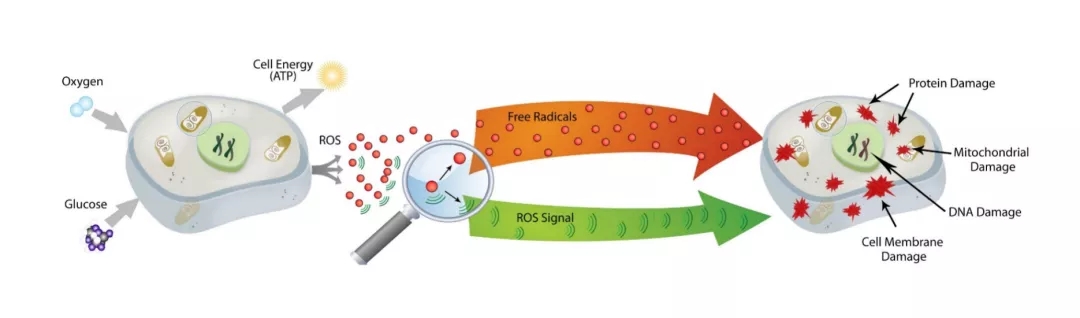
线粒体与 ROS 信号传导
线粒体直接利用氧气制造能量,而大量电子总会在高氧环境和高还原态的呼吸链中泄漏,产生多种氧化磷酸化的天然副产物——氧的单电子还原产物,如超氧阴离子、单线态氧、过氧化物等,统称活性氧(ROS)。过量ROS与自由基泄漏到线粒体外进入细胞及胞外,将电子传递给其他分子,对人体造成的氧化损害,称为氧化应激。事实上,线粒体正常代谢产生的适量ROS作为信号分子具有重要生理作用,而过量摄入外源性抗氧化剂可造成机体对ROS信号不敏感,导致低效线粒体无法及时清除,反而引发ATP长期不足,加速衰老。因此,减少氧化损害更重要的是减少自由基的内源性产生,提升线粒体自身抗氧化能力,而非一味补充外源性抗氧化剂。
mtDNA突变与衰老
mtDNA突变是皮肤衰老,尤其是皮肤光老化的潜在根本原因。随着年龄增长,mtDNA突变会在复制后的组织中积累,伴随线粒体功能下降、ROS产生增加以及线粒体膜电位丧失,继而导致线粒体自噬和细胞凋亡增强。与皮肤mtDNA突变相关的外界因素包括日晒、吸烟、环境及食物中的化学因素、X射线与化学性致癌剂等。mtDNA突变后所产生的生物学效应有两种模式:细胞生物能阈值模式和细胞程序性死亡模式。发生在mtDNA结构基因上的突变大多与细胞呼吸功能有关,一方面造成线粒体呼吸相关功能组分的合成障碍和缺失,导致线粒体呼吸能力下降,引发细胞衰老;另一方面造成mtDNA复制与修复障碍,加重线粒体损伤,加剧细胞衰老。最新研究表明mtDNA是皮肤衰老起皱、脱发的可逆调节因子,而恢复线粒体功能可以使皮肤恢复年轻。
线粒体与炎症及免疫调控
线粒体,特别是mtDNA,在机体炎症和免疫调控中发挥核心作用。氧化应激是炎症反应的重要引发机制,而线粒体是ROS生成的主要场所。mtDNA作为细胞损伤相关分子模式(DAMPs)之一,是线粒体损伤与炎症功能间联系的标志物。一方面,mtDNA通过激活三条炎症通路的相应受体,即Toll样受体蛋白(TLR)、核苷酸结合寡聚化结构域样受体蛋白(NLRP)和细胞质循环GMP-AMP合成酶-干扰素基因刺激因子DNA(cGAS-STING DNA)传感系统介导的途径参与炎症反应,同时损伤线粒体释放甲酰多肽激起免疫应答;另一方面,线粒体功能与ROS代谢异常同样可以激发炎症反应。过量ROS会降解细胞膜的磷脂分子,在磷脂酶A2的作用下诱发花生四烯酸,合成初级炎症因子白三烯(LTB4)和前列腺素(PGE2),导致红斑、肿胀、致热效应等肌体炎症;同时过量ROS也会导致线粒体脂质过氧化、膜电位崩溃和ATP合成障碍,进一步诱发炎症。线粒体既是ROS的主要来源,又是其损伤靶点,两者相互作用,共同调节炎症进程。持续炎症刺激可激活循环免疫细胞,循环释放细胞因子、趋化因子、一氧化氮(NO)和ROS损伤线粒体,反向激活mtDNA诱导炎症通路,形成恶性循环。
此外,衰老的标志之一是低度、慢性、无菌炎症状态的发展,通常被称为“炎性衰老(Inflammaging)”,而线粒体功能与年龄依赖性免疫反应的增强激活密切相关。衰老细胞会产生一系列衰老相关分泌表型(SASP),包括基质金属蛋白酶(MMP)、生长因子(GF)和促炎症细胞因子。SASP的形成与线粒体损伤调节的免疫应答的激活过程密切相关,可加重衰老细胞的促衰老效应。线粒体的代谢、稳态改变、自嗜、凋亡过程均与炎性体的激活密切相关,影响细胞及肌肤老化进程。
线粒体与
损伤修复、胶原合成
线粒体全面参与损伤修复的炎症期、增殖期、塑形期三个阶段。线粒体一方面提供细胞生长所需ATP能量基础;另一方面提供氧气与营养等物质基础,促进毛细血管新生,改善微环境,清除坏死组织。同时,线粒体内储存的Ca2+能上调一氧化氮合酶(NOS)活性,诱发血管新生,加速基质重塑,并诱导成纤维细胞、内皮细胞等增殖。此外,线粒体参与调控VEGF、bFGF和PDGF等生长因子释放,进而促进血管内皮分裂与血管再生,促进肉芽生成与损伤修复。同时,线粒体膜电位皮肤呈纤维细胞状态,功能障碍的线粒体膜电位发生去极化,导致成纤维细胞功能过度活跃、ATP水平显著升高及过度增殖,产生过量胶原蛋白,进而引起瘢痕异常增生,影响肌肤修复。
线粒体动力学:融合与分裂
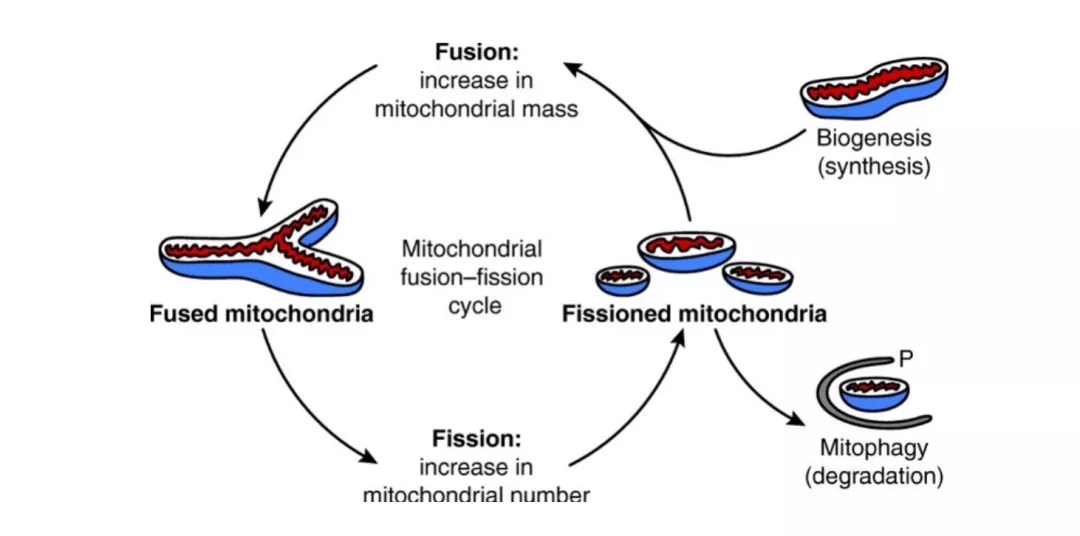
线粒体动力学
线粒体实际上是一个互联系统,通过融合和分裂动态调节自身形态和功能,维持mtDNA完整性、电和生化连通性,以及周转、分离或保护。其动态平衡有助于线粒体质量控制——融合允许在完整的和功能异常的线粒体之间进行信号传递和内容物交换等相互联系,替换受损的mtDNA,以促进线粒体完整性,并在细胞内提供质量均匀的线粒体群体;分裂允许分离不可逆损伤及功能缺陷的线粒体,并通过线粒体自噬或程序性细胞死亡过程将其清除。
线粒体自噬与细胞凋亡
自噬(Autophagy)和线粒体自噬(Mitophagy)在表皮角质形成细胞分化中起关键作用,通过选择性清除衰老、损伤及功能失调的线粒体,有助于降低细胞内ROS,缓解细胞衰老表型,从而保持细胞活性。线粒体自噬与炎症息息相关,在致炎因子刺激下,线粒体自噬水平升高,可清除损伤线粒体,并促进健康线粒体增殖来改善线粒体稳态和功能,发挥抗炎效应。此外,细胞自噬以“自噬体”形成为特征,宿主借助线粒体清除损伤细胞器、蛋白降解物及废弃细胞。
细胞凋亡是一系列以凋亡蛋白酶(Caspases)激活为基础、以不引起炎性反应为特征的病理过程。2014年,Rongvaux与White同时独立发现线粒体是决定死亡体细胞“命运”的核心因素:一方面死亡的体细胞释放mtDNA,经cGAS-STING通路生成一系列炎性因子;另一方面线粒体内Caspases可自发激活,抑制DAMPs(尤其是mtDNA)引起的炎性反应。线粒体这种“双向”功效在维持宿主免疫平衡中至关重要。Caspase激活的内在途径由线粒体调控,线粒体外膜通透性(MOMP)导致细胞色素c的释放是主要的凋亡信号;反过来,MOMP受线粒体内Bcl-2蛋白质家族控制。增强线粒体膜通透性屏障可以增强表皮角质形成细胞对日晒损伤的防护。
细胞凋亡是一系列以凋亡蛋白酶(Caspases)激活为基础、以不引起炎性反应为特征的病理过程。2014年,Rongvaux与White同时独立发现线粒体是决定死亡体细胞“命运”的核心因素:一方面死亡的体细胞释放mtDNA,经cGAS-STING通路生成一系列炎性因子;另一方面线粒体内Caspases可自发激活,抑制DAMPs(尤其是mtDNA)引起的炎性反应。线粒体这种“双向”功效在维持宿主免疫平衡中至关重要。Caspase激活的内在途径由线粒体调控,线粒体外膜通透性(MOMP)导致细胞色素c的释放是主要的凋亡信号;反过来,MOMP受线粒体内Bcl-2蛋白质家族控制。增强线粒体膜通透性屏障可以增强表皮角质形成细胞对日晒损伤的防护。
线粒体与干细胞
干细胞对皮肤自我修复和再生至关重要,干细胞衰老与线粒体功能衰退密切相关。线粒体产生的ROS对干细胞增殖分化有调节作用,并通过调节代谢物的比率调节干细胞自我更新和稳态。同时,线粒体可通过生物合成和融合分裂等形态结构的动态平衡维持干细胞多能性、合成代谢、氧化还原平衡、诱导分化及干细胞重编程。此外,线粒体自噬可以有效清除干细胞中功能缺陷的线粒体,维持干细胞功能。干细胞的独特性质表明其有机制确保自身不会积累陈旧和功能障碍的线粒体。基于此,Lonergan等提出将线粒体形态、定位、数量及功能作为评价干细胞是否具备多能性的标志。此外,还存在“线粒体转移”现象,即干细胞将其正常线粒体转移到线粒体功能障碍的应激细胞中,重建呼吸功能和ATP供能。
线粒体与长寿
在百岁老人中,遗传性线粒体DNA单倍群与长寿相关;而患有线粒体遗传疾病的个体表现出早衰表型。已知卡路里限制可以延缓寿命,这最初被解释为因其可以减慢糖代谢、呼吸作用及ROS产生;然而与直觉相反,现已知这是长寿蛋白Sirtuin(其中三种——3、4和5位于线粒体)等通过线粒体自噬等途径上调其功能的结果。而仅存在于线粒体中的长寿基因SIRT3的高活性可作为长寿的标志。
线粒体与钙稳态
线粒体是细胞内重要的Ca2+池,线粒体呼吸需要摄取Ca2+,并且参与控制Ca2+浓度、皮肤细胞分裂分化、皮脂合成分泌、皮肤屏障功能再生、敏感肌修复以及预防医美术后返黑等过程。线粒体通过线粒体钙离子单向转运蛋白(MCU)、雷诺丁受体(RyR)等摄取Ca2+,通过钠钙交换体(NCX)、线粒体渗透性转换孔(mPTP)等释放Ca2+,调节自身与胞内Ca2+浓度,维持Ca2+稳态,调节细胞凋亡与自噬。ROS可引起线粒体内外Ca2+稳态失衡,使细胞外Ca2+大量内流,产生“钙超载”,导致线粒体、细胞、组织功能障碍。过量Ca2+还可以激活诱导性一氧化氮合酶、ROS和NO生成增多。
线粒体与紫外损伤及光老化
光老化皮肤的表型特征是线粒体功能障碍和细胞外基质降解的结果。其引发机制包括ROS和氧化应激标志物水平升高、DNA损伤、mtDNA突变、细胞凋亡、MMPs相关胶原蛋白作用、炎症反应及免疫抑制、晚期糖基化终末产物(AGEs)作用等,其作用过程均与线粒体功能直接相关。UVA构成UV大部分,是光老化的主要参与者,可以更深地穿透到真皮中,但仅间接影响DNA;UVB主要作用于表皮角质形成细胞和黑色素细胞,但会直接造成DNA损伤。UVB一方面通过DNA直接损伤、诱导ROS形成及活化细胞膜表面死亡受体等方式引起角质形成细胞凋亡;另一方面激活炎症信号传导机制,引起肌肤免疫和炎症反应。成纤维细胞位于表皮下的真皮层,却更易发生UV及IR诱导的氧化损伤。UV诱导的光老化与mtDNA缺失直接相关。据研究,光老化皮肤细胞mtDNA缺失率比防晒保护的皮肤细胞高10倍。
线粒体与黑色素及
褪黑素-线粒体轴
线粒体功能是皮肤色素沉着的调节剂,其功能障碍直接或间接通过过量ROS信号和褪黑素生成而干扰黑色素的生物合成。黑色素生成具有助氧化作用,且较其他细胞ROS基础水平更高,因此黑色素细胞极易受到氧化损伤。位于线粒体内膜中的抑制素蛋白可直接与黑色素生成素结合。黑色素的光降解可能是光暴露皮肤所表现出的氧化应激和mtDNA损伤的重要机制。
皮肤细胞中的褪黑素除了调节睡眠节律、伤口愈合、黑色素生成、色素沉着、炎症和免疫反应外,还具有抗氧化和光保护作用,其代谢严重依赖线粒体和ROS信号传导。褪黑素与表皮线粒体之间存在“共生”关系——线粒体是皮肤褪黑素生物合成和代谢的场所,而褪黑素对线粒体ETC和抗氧化机制具有调节作用。除了直接清除诸如H2O2、单线态氧和NO-之类的ROS外,褪黑素及其衍生物还可以通过增加抗氧化酶的活性、介导细胞色素c的还原来保护皮肤细胞免受辐射损伤。此外,褪黑素也与细胞内调节Ca2+稳态的钙调蛋白结合,并可通过增加角质形成细胞的增殖来改善表皮的屏障功能,还可加速伤口愈合,增强抗菌药物的效果。
线粒体与污染
空气污染对皮肤的损害日益受到关注。PM2.5和臭氧可通过线粒体ROS和氧化应激来增加DNA和蛋白质损伤,并通过影响色素沉着和皱纹与皮肤衰老呈正相关。与交通相关的颗粒和烟灰与前额及脸颊色素斑点的形成有关,表明UV辐射和污染应激源之间存在共同机制,并可能源于线粒体复合体II的年龄依赖性下降。色素过度沉着会导致复合体II的活性抑制和超氧化物ROS增加,同时黑色素浓度的增加能减弱其进一步形成。因此色斑的形成可能是防止衰老皮肤细胞产生更多ROS的保护机制,并可依此开发靶向线粒体的抗污染方案。
线粒体与脱发白发
脱发和白发是可观察到的关键衰老表型。有证据支持线粒体通过增加氧化应激(为主)和降低自身容量(为辅)这两条途径参与其中。表皮和毛囊上皮中mtDNA的缺失和电子传递链复合物的丢失,会表现出黑色素生成和毛发生长的功能异常。来自线粒体功能的ROS信号在毛囊形态发生和毛干伸长中起关键作用。人类与年龄相关的脱发主要是雄激素性的,且男性发病率更高。秃顶区域的真皮乳头细胞会表观出更高水平的线粒体相关衰老标记物。而mtDNA修复机制故障导致的mtDNA高突变丰度和加速的衰老表型,会表现为头发密度的减小。
白发的特征是色素单元中的黑色素细胞达到了最高水平的氧化应激,因凋亡而逐渐丧失,并且使线粒体凋亡途径的抑制剂Bcl-2丧失了清除ROS的活性,直到毛囊失去色素。
线粒体护肤成分
尽管美护行业中存在一些可以促进生成ATP或改善线粒体其他功能的植物提取物或天然化合物,然而以下常见活性物质的功效更为专业人士认可。
烟酰胺,也称为维生素B3,是线粒体生成ATP时所必需的NAD+的前体,可上调Sirtuins,通过线粒体自噬提高线粒体质量,发挥多样化的生理功能。 辅酶Q10直接存在于线粒体电子传递链中,在复合体I/II和复合体III之间起电子穿梭作用,可以恢复ATP产生,抑制线粒体去极化,防止线粒体自噬,并作为防止膜脂氧化的ROS清除剂,是重要的抗氧化剂和非特异性免疫增强剂。 白藜芦醇可通过增加细胞器生物合成和减少ROS产生,尤其是上调Sirtuins中的SIRT1,直接改善线粒体功能,发挥抗氧化及抗衰老作用。 麦角硫因已被证实可通过转运蛋白OCTN1在线粒体中发挥独特的抗氧化功能,清除自由基,从而抑制DNA损伤,减少脂质过氧化,防护UV损伤。 此外,肌肽因其良好的线粒体ROS淬灭能力,以及对线粒体过氧化物酶、脂



关闭




扫码联系
扫码关注








